
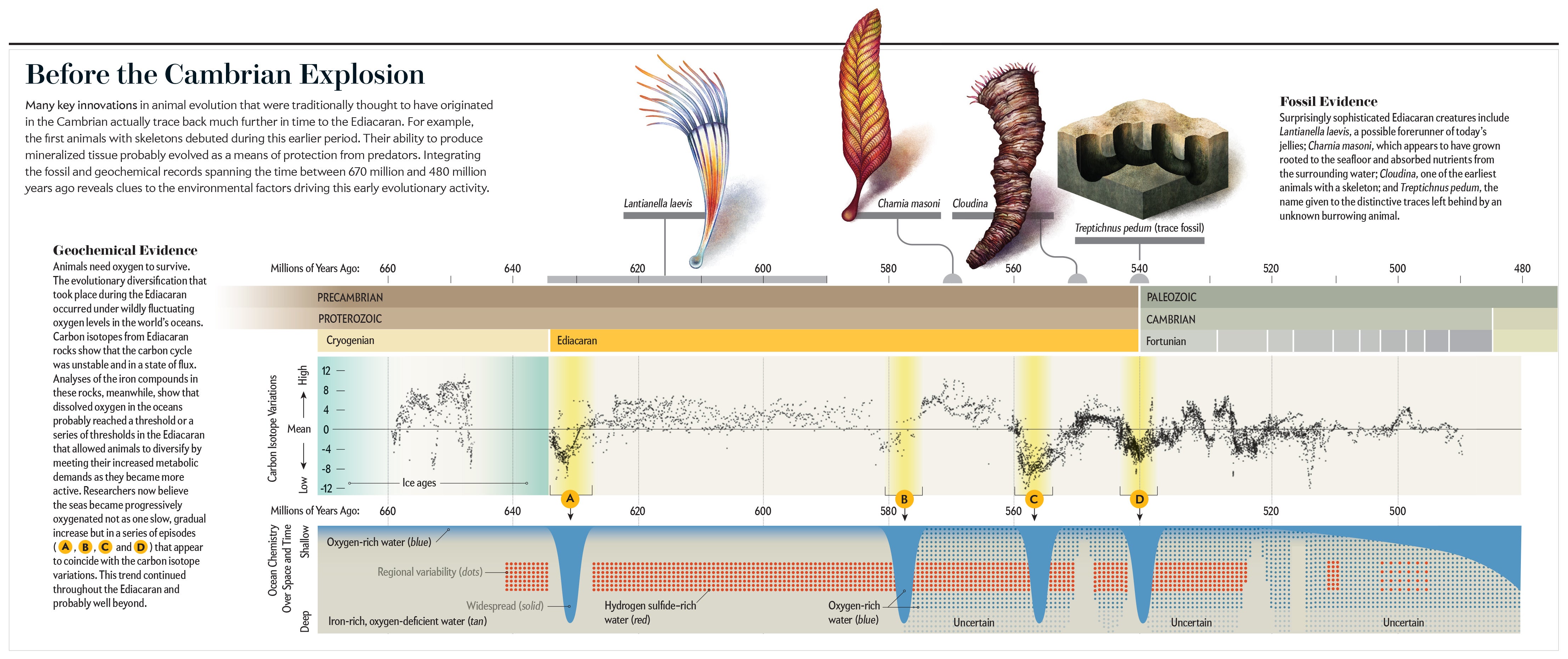
站在環繞西伯利亞巨河的陡峭白色懸崖頂上,您的腳下將標誌著地球生命史上一個關鍵點:前寒武紀和寒武紀時期之間有5.41億年曆史的地質分界線。這條分界線以下的岩石包含稀少的化石遺蹟——軟體生物的幽靈般的印痕和少量的殼狀結構。但是,敲開分界線以上的任何岩石,它們都將佈滿貝殼。再往上一點,就會出現熟悉的化石生物,例如三葉蟲。這些變化記錄了所謂的寒武紀大爆發,這是進化史上最重要但仍然知之甚少的事件之一。
幾十年來,科學家們一直認為複雜的動物——具有分化組織型別的多細胞生物——起源於寒武紀大爆發。可以肯定的是,在此期間,包括當今許多主要動物群體的祖先在內的大量新穎形態爆發式出現。但是,來自西伯利亞、奈米比亞和其他地方的最新發現表明,複雜的動物實際上在寒武紀大爆發之前的數百萬年,在前寒武紀的最後一個章節——埃迪卡拉紀期間就開始出現。在這些發現中,已知最古老的生物具有由礦化組織構成的外部和內部骨骼,這是許多現代動物身上可見的關鍵進化創新。
這些帶甲生物在如此久遠的時間——5.5億年前——的存在表明,人們認為驅動寒武紀大爆發的生態和環境壓力實際上在那之前很久就已經在起作用了。弄清楚這些因素如何在埃迪卡拉紀塑造了最早的複雜動物的進化,是理解隨後在寒武紀發生的驚人多樣化爆發的關鍵。
支援科學新聞報道
如果您喜歡這篇文章,請考慮透過以下方式支援我們屢獲殊榮的新聞報道 訂閱。透過購買訂閱,您將幫助確保有關塑造我們當今世界的發現和想法的有影響力的故事的未來。
寒武紀化石記錄一直是150多年來深入研究的主題。因此,寒武紀化石在全球範圍內出現的時間和地點的總體模式已相對確定:在許多大陸的大約同一時間發現了相似的化石,並且它們或多或少同步地遵循相同的進化變化序列。但是,直到現在,隨著更古老的埃迪卡拉紀化石的發現,我們才開始看到寒武紀大爆發的根源。
令人欣慰的是,我們也開始弄清楚它為何在當時發生,這部分歸功於新的地球化學技術的開發,這些技術徹底改變了我們對埃迪卡拉紀-寒武紀世界海洋化學變化的理解。來自新興的化石和地球化學記錄的見解最近才被整合起來,以展示地球的生物圈、岩石圈、水圈和大氣圈——統稱為地球系統——在此期間可能如何運作。但是,我們已經可以描繪出一幅引人注目的圖景,展示了海底如何在寒武紀大爆發之前的數千萬年中,逐漸被越來越複雜的生物所佔據,為我們所知的動物生命的興起奠定了基礎。
首批動物
古代動物最古老的可能證據並非來自可識別的化石,而是來自被稱為生物標誌物的有機化合物的遺骸。研究人員在阿曼胡克夫超群 (Huqf Supergroup) 的儲存非常完好的岩石中發現了一種這樣的生物標誌物,一種特定形式的甾烷,其歷史至少有6.5億年。一些專家認為,這些甾烷是特定於一組海綿的,因此,這些分子在胡克夫岩石中的存在證明了這些動物在那個非常早期的時代就已存在。然而,並非所有科學家都接受這些甾烷是海綿特有的說法。事實上,四月份發表的一項研究表明,它們是單細胞變形蟲群體的診斷特徵。
最古老的候選動物化石來自中國南方一個名為藍田組的岩層序列,可能早至6.35億年前,同樣存在爭議。一些研究人員認為,這些微小的軟體生物與珊瑚或水母有關,因為它們表現出觸手狀結構,但是這些化石的儲存狀況不夠清晰,無法做出明確的解釋,這使得許多研究人員不相信它們代表任何型別的動物。

化石蹤跡:複雜動物的關鍵化石來自西伯利亞尤多馬河岸邊(上圖)和奈米比亞奈米布沙漠邊緣(下圖)的埃迪卡拉紀岩石。圖片來源:瑞秋·A·伍德
幾乎所有人都能認同的最古老動物遺骸是來自紐芬蘭的化石,其年代可追溯到約5.71億年前,就在最後一次區域性“雪球地球”冰川期之後,那次冰川期將地球大部分地區包裹在厚厚的冰層中。這些最早已知的埃迪卡拉生物群的代表主要是高度或寬度達一米的軟體生物。有些呈現出大型羽毛狀葉狀體的形式,帶有垂直莖,將其紮根於海底;另一些則蔓延在海底,其扁平的身體呈現出分形結構,分支單元在所有尺度上都顯示出相同的模式。所有這些身體構造都最大化了表面積,表明這些動物直接從周圍的水中吸收營養。
這種適度多樣的動物群持續了1000多萬年。但是隨後,動物進化的步伐開始加快。化石記錄表明,大約在5.6億年前之後,埃迪卡拉生物群開始多樣化,包括棲息在淺海中的移動形態。一些化石保留了刮擦痕跡,表明這些動物透過啃食藻類墊來攝食。另一些可能拖著身體穿過藻類,從身體底部吸收營養。最早的簡單洞穴也大約在這個時候出現,這證明動物已經開始移動並擾亂海底的沉積物。
快進到大約5.5億年前,在石灰岩岩石(主要由碳酸鈣組成)中突然出現了儲存外部和內部骨骼的最古老化石。這些化石在大小和形態上已經多種多樣,並且出現在西伯利亞、巴西和奈米比亞等遙遠的地方。世界各地如此多不相關的動物群體在此時出現骨骼,證明了在全球範圍內起作用的主要驅動進化力量。我們不確定這種力量是什麼。但是我們有一個想法。製造骨骼在能量上是昂貴的,因此對於動物來說,要進行這種努力,收益必須大於成本。動物可能會出於多種原因產生骨骼,但到目前為止,最常見的原因是需要保護自己免受捕食者的侵害。儘管此時期沒有捕食者的化石證據,但有理由認為骨骼的出現可能反映了首次廣泛出現的以其他動物為食的動物。
同舟共濟
最近對這些古代骨骼的分析為了解其所有者的外觀以及它們的生活方式提供了誘人的線索。一種名為Cloudina的生物,因其精緻的管狀骨骼化石而聞名,該骨骼的長度可達約70毫米,類似於一堆冰淇淋蛋筒,在我們的埃迪卡拉紀生態系統重建中發揮了重要作用。Cloudina於1972年在奈米比亞首次被發現,長期以來一直被認為生長在海底。但是在過去的幾年中,研究人員從世界各地的地點發現了許多新的Cloudina標本,這些標本改變了這一觀點。我的團隊對來自奈米比亞的標本的研究表明,Cloudina具有多種生長方式。它可以附著在由微生物製成的墊子上,這些墊子結合了海底的軟沉積物,或者它可以將自身錨定在藍藻的分層土墩上。最重要的是,Cloudina個體實際上可以將自己粘合在一起以形成珊瑚礁。這一發現確立了Cloudina是最古老的造礁動物之一,將這種生活方式的記錄推後了大約2000萬年。
Cloudina是否與現代造礁珊瑚(如珊瑚)有關仍然不確定。但是我們確實知道,像造礁珊瑚一樣,它與許多其他動物生活在一起。這種密切聯絡的線索來自在與含有Cloudina化石的岩石相同年齡的岩石中發現的其他骨骼化石。一種名為Namacalathus的生物,從世界各地的化石產地得知,似乎是Cloudina的夥伴之一。它的骨骼長達50毫米,由細而薄壁的莖和一個杯狀結構組成,頂部有一箇中央開口,側面有幾個開口。動物的軟組織可能主要在杯狀結構內部,儘管它從未被儲存下來。Namacalathus的化石表明它生長在微生物墊上,通常靠近Cloudina。
圖片來源:弗朗茨·安東尼(Franz Anthony)(化石);來源:《環境變化和進化的綜合記錄挑戰寒武紀大爆發》,瑞秋·伍德 (Rachel Wood) 等人,《自然生態與進化》雜誌,第3卷;2019年4月
Namapoikia是一種僅從奈米比亞化石產地得知的生物,也與Cloudina為伍。這種動物以其巨大的尺寸——直徑達一米——和堅固的骨骼而聞名。根據其生長形態,我們認為Namapoikia是一種海綿,因此應該具有內部骨骼,這與Cloudina和Namacalathus可能具有的外部骨骼形成對比。有趣的是,Namapoikia生長在珊瑚礁的隱蔽處,覆蓋著開放裂縫和裂隙的垂直壁。在現代珊瑚礁中,生活在開放表面的動植物群落與佔據洞穴、縫隙或下垂物等更隱蔽區域的動植物群落不同。我們的埃迪卡拉紀化石發現表明,這種區別與動物珊瑚礁本身一樣古老。
這些觀察結果意義重大,因為造礁代表著重要的生態創新。透過緊密地生長在一起甚至粘合在一起,個體可以變得機械強度更高,從海底升起遠離競爭對手,提高攝食效率並獲得免受捕食者侵害的保護。因此,像最早的骨骼一樣,埃迪卡拉紀化石記錄中珊瑚礁的出現可能預示著不斷上升的複雜生態壓力。寒武紀大爆發,以及捕食者和獵物之間的軍備競賽,早已開始。
埃迪卡拉紀世界
到2010年代中期,人們越來越清楚地認識到,寒武紀並沒有標誌著專家長期設想的與埃迪卡拉紀的突然、戲劇性的決裂。研究人員不僅開始積累證據表明動物開始進化骨骼和建造珊瑚礁的時間早於傳統認為的時間,而且我們還開發了生態系統模型,表明埃迪卡拉紀動物群落與寒武紀動物群落具有許多生態特徵。“大爆發”,我們正在瞭解到,其導火索比以前認識到的要長得多。
然後,在幾年前,西伯利亞和中國的一些關鍵發現進一步模糊了埃迪卡拉紀和寒武紀世界的界限。來自中國和德國的一組研究人員發現,Cloudina一直延續到寒武紀。我的團隊與來自俄羅斯和中國的同事一起,在埃迪卡拉紀岩石中發現了長期以來被認為是寒武紀特有的化石。這些發現向我們強調,要解開寒武紀大爆發的謎團,我們必須弄清楚這些動物起源的埃迪卡拉紀世界的動態。
氧氣供應變化可能起到的作用是研究人員特別渴望確定的一個動態因素。動物需要氧氣,因此過去幾年來的一箇中心辯論是瞭解在埃迪卡拉紀和寒武紀之間的時間跨度內的某個時刻,氧氣水平是否上升超過某個臨界閾值,從而使動物能夠蓬勃發展。這個問題比看起來更復雜,因為動物並非都具有相同的氧氣需求。簡單、不動的生物,例如海綿,可能比移動動物需要更少的氧氣,並且它們當然比活躍、快速遊動的捕食者所需的氧氣少得多。在我們的調查過程中,我們牢記了這種變化。

石刻文字:Cloudina是最古老的具有外部骨骼的生物之一,從儲存其精緻管狀裝甲的化石中得知(左上)。Cloudina個體可以將自己粘合在一起,形成珊瑚礁。另一種早期的骨骼形式是Namacalathus,其杯狀硬質部分通常大量儲存(右上)。Cloudina和Namacalathus經常一起發現(左下)。Namapoikia,一種早期海綿,是Cloudina的另一種夥伴,它生長在珊瑚礁的隱蔽縫隙中(右下)。圖片來源:瑞秋·A·伍德
對我們來說幸運的是,近年來開發了許多新的地球化學方法來估計這些古代海洋中存在的氧氣量。一種特別強大的技術——鐵形態分析——利用了各種鐵化合物的特性,這些化合物的行為方式取決於是否存在氧氣。這種方法使我們能夠在區域性範圍內看到在何處——以及何時——有足夠的氧氣來支援複雜的生命。使用這種方法進行的研究所得出的廣泛共識是:海洋中的溶解氧可能在埃迪卡拉紀期間達到了一個或一系列閾值,從而使動物能夠透過滿足其日益增長的代謝需求而多樣化,因為它們變得更加移動和活躍。
科學家們現在已經組裝了足夠大的地球化學資料集,我們可以重建氧氣不僅在特定年齡的個別埃迪卡拉紀地點如何分佈,而且在全球範圍內隨時間如何分佈。這項工作揭示了整個埃迪卡拉紀和早期寒武紀的模式,這些模式與今天的模式大相徑庭,許多地區顯示出相對薄的一層富氧淺水層,覆蓋著可能完全缺乏氧氣的較厚楔形深海水,這種狀態被稱為缺氧。
這些地球化學資料還表明,缺氧水和富氧水之間的邊界在此期間非常活躍,隨著海平面變化而上升和下降。因此,早期動物可居住的淺海海底區域比科學家預期的還要受限——名副其實的富氧水綠洲。如果埃迪卡拉紀和寒武紀期間發生的進化多樣化是在相對較低的氧氣水平下但在高度動態的條件下發生的,這些條件在生態、全球和進化時間尺度上波動,那麼這些因素可能如何塑造了那次非凡的輻射?
創新的引擎?
海底缺氧時期與一些著名的物種大滅絕相吻合,例如2.52億年前的二疊紀大滅絕,導致超過90%的海洋物種滅絕。但是,幾次主要的物種多樣化——包括埃迪卡拉紀-寒武紀、大約1億年後的奧陶紀和大約2.47億年前的中晚三疊紀——都始於長期動態淺海缺氧時期。考慮到這些事件,我的同事史密森學會的道格·歐文 (Doug Erwin) 和我假設,波動的氧氣條件可能為軟體動物的進化創新創造了關鍵機會。
當海水氧氣水平超過每升10微摩爾時,動物更容易形成石灰岩骨骼——石灰岩是許多現代海洋生物的骨骼和貝殼的材料。也許軟體動物只有在氧氣水平達到這樣的閾值後才能進化出這些碳酸鈣骨骼,從而使以前孤立的綠洲能夠在全球範圍內擴張、連線並實現穩定。
關於生命可能如何在進化時間尺度上響應氧氣可用性的變化,還有很多有待發現。這種響應可能很複雜,因為動物還在與捕食增加等其他因素作鬥爭。而且,由於個體生物、生態系統和更廣泛的地球系統之間的反饋(這些反饋在很大程度上是未知的)也將被納入方程。
我們還有很多工作要做。在整個埃迪卡拉紀-寒武紀期間塑造地球地殼的區域過程的劇烈變化,在地質和化石記錄中產生了許多重大空白。這意味著我們必須從世界各地眾多地點收集的資料中拼湊出我們關於複雜動物興起的敘述。許多關鍵的埃迪卡拉紀地點的年代仍然很差,這進一步複雜化了我們的任務。我們通常透過測量古代火山噴發產生的附近火山灰層中鋯石晶體中鉛與鈾的比率來確定這個年代的岩石的年代。這是少數幾種可以為給定岩石提供絕對放射性年代的方法之一。但是令人沮喪的是,我們許多最著名的層序都缺乏這些重要的火山灰層。結果,我們無法準確地關聯世界不同地區發生的進化變化,這對於為我們的事件歷史建立堅實的時間框架至關重要。一個主要的例子是中國備受爭議的藍田組,該組已產生最古老的候選動物化石,但其年代可能在6.35億年至5.9億年之間的任何位置。
儘管如此,仍有理由保持樂觀。新的火山灰層正在出現,年代測定方法正在改進。例如,許多研究小組用於計算在奈米比亞發現的埃迪卡拉紀化石年代的火山灰層最近已被重新測定年代,最年輕的火山灰層——那些最靠近前寒武紀-寒武紀邊界的火山灰層——已被證明比以前認為的年輕兩百多萬年。這一結果引發了關於這些化石實際上如何與紐芬蘭和西伯利亞以及其他關鍵地點的同類化石相關的重大問題。此外,地球化學家正在開發新的同位素技術和其他方法,這些方法可以使我們對這個古代世界的氧氣狀況的認識更加清晰。我的團隊和其他人正在人跡罕至的地方(例如西伯利亞)尋找新的化石,這些地方在很大程度上仍未被探索。
在不遠的將來,當我們站在那些懸崖上,俯瞰下方廣闊的森林時,我們將對這段最非凡的時間有更深入的瞭解。