
我們的記憶取決於我們回憶關於世界的細節的能力——孩子的臉龐、一隻鵝、一個湖泊。然而,為了將它們轉化為真實的體驗,大腦必須以某種方式將這些獨立的元素融合成一個完整的整體——當她看到一群鵝突然從湖畔的蘆葦叢中飛起時,孩子臉上的表情。
連貫的記憶感也依賴於其他因素。我們數千年的生存依賴於不僅回憶起正確的資訊——比如,獅子或蛇——還要回憶起背景。我們是在非洲稀樹草原的孤立地帶遭遇了一場突如其來的動物遭遇戰,還是在聖地亞哥動物園悠閒地觀看?
為了避開我們日常生活中的其他型別的捕食者,我們也需要能夠隨著時間的推移連線記憶:判斷一項看似有吸引力的投資是否值得追求,取決於推薦來源——例如,建議者的可靠性。未能將兩者聯絡起來可能會導致災難性的後果。
支援科學新聞報道
如果您喜歡這篇文章,請考慮透過以下方式支援我們屢獲殊榮的新聞報道 訂閱。透過購買訂閱,您正在幫助確保有關塑造我們當今世界的發現和思想的有影響力的故事的未來。
神經科學領域開始努力解決大腦如何在空間和時間上連線記憶。到目前為止,絕大多數研究都集中在我們獲取、儲存、回憶和改變單個記憶的方式上。然而,大多數記憶並非作為單一、孤立的實體而獨立存在。相反,一個回憶會召喚下一個回憶,建立起復雜的記憶序列,幫助我們更好地預測和理解我們周圍的世界。
大腦用來創造這些關聯記憶的基本機制正開始顯現出來——這得益於我的實驗室和其他實驗室 20 年的研究。理解交織個體記憶所涉及的物理過程,不僅可以深入瞭解大腦的工作方式,還可以幫助我們預防破壞我們創造和聯絡記憶能力的記憶障礙。
意外的驚喜
當我們在 20 世紀 90 年代後期開始研究記憶關聯時,我們缺乏解決這個問題所需的工具和基本知識。確定記憶如何相互交織的關鍵第一步是我們發現了一個稱為記憶分配的概念,即大腦使用特定規則將學習資訊的片段分配給大腦區域中參與形成記憶的離散神經元群體的認識。
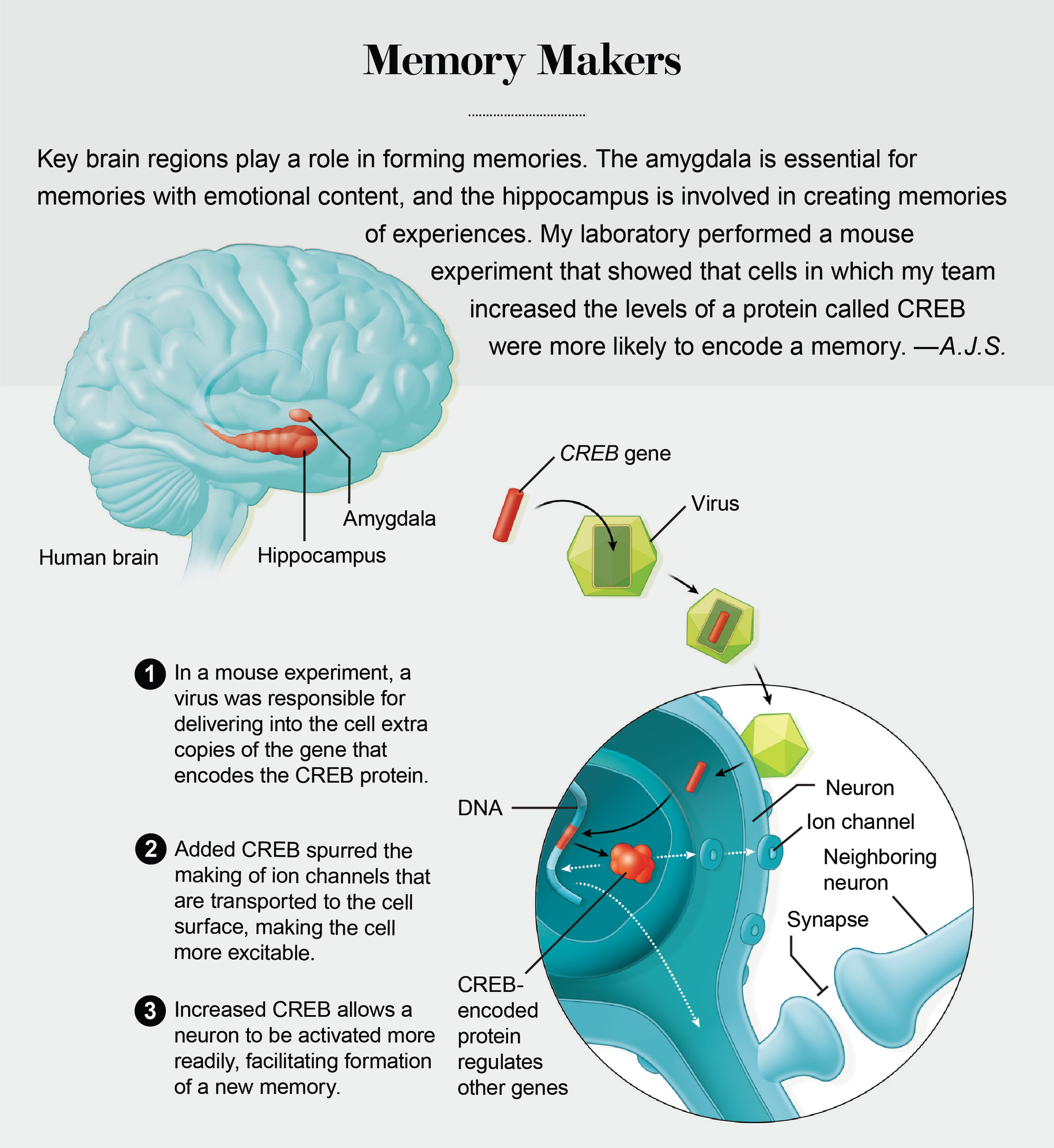
巧合在記憶分配的發現中發揮了關鍵作用。這始於 1998 年我在耶魯大學訪問期間與朋友兼同事、現任埃默裡大學的邁克爾·戴維斯的一次談話。戴維斯與我分享了他的實驗室研究的發現,其中他的實驗室操縱了一種稱為CREB的基因,以增強大鼠的情緒記憶——例如,音調與電擊之間的關聯。此前,我的實驗室(現在位於加州大學洛杉磯分校)和其他研究人員已經表明,形成長期記憶需要CREB基因。CREB 基因透過編碼一種調節記憶所需其他基因表達的蛋白質來完成這項任務。在學習過程中,一些突觸(神經元用來交流的細胞結構)會被建立或加強,以便它們可以促進細胞之間的相互作用。CREB 蛋白質充當這個過程的分子架構師。沒有它的幫助,大多數經歷都會被遺忘。
令我驚訝的是,戴維斯的研究小組能夠改善記憶,即使他的實驗室僅在杏仁核(大腦中對情緒記憶至關重要的區域)的整體神經元群體的一個子集中提高了 CREB 水平。在我訪問耶魯大學後的幾個月裡,一直困擾我的問題是,記憶是如何最終進入到少數可以利用更高 CREB 水平的細胞中的?難道 CREB 不僅協調了記憶的形成,還有助於確保具有 CREB 的細胞更有可能參與記憶的形成嗎?在我們自己對 CREB 的研究中,我們將重點放在了我們已知與記憶相關的特定大腦區域內的功能:杏仁核和海馬體。後者儲存了一個人周圍環境的內部地圖。

顯微鏡安裝在活體小鼠的頭部,讓研究人員檢查儲存記憶的大腦細胞的活動。圖片來源:Denise J. Cai 加州大學洛杉磯分校整合學習與記憶中心
科學就像尋找問題一樣,也像回答問題一樣。與戴維斯的談話幫助我意識到,神經科學家對給定記憶如何分配到處理和儲存我們回憶的大腦區域的神經元中的規則知之甚少,如果真的有規則的話。因此,我們決定更仔細地觀察。
我們的第一個重大突破是在我們招募了神經科學家希娜·A·喬斯林之後,她曾在戴維斯實驗室研究 CREB。在她在我的實驗室以及後來在多倫多大學的她自己的實驗室與同事進行的一系列動物實驗中,喬斯林使用病毒將 CREB 的額外複製引入小鼠杏仁核內的特定神經元。她表明,與鄰近的神經元相比,這些神經元儲存恐懼記憶的可能性幾乎高出四倍。
2007 年,經過近十年的努力,我的實驗室與喬斯林的團隊合作,最終發表了證據,表明情緒記憶並非隨機分配到杏仁核內的神經元。相反,被選來儲存這些記憶的細胞是那些具有更多 CREB 蛋白質的細胞。同樣重要的是,隨後的實驗表明,CREB 在其他大腦區域(包括海馬體和大腦皮層,即最外層)也具有類似的功能。
開啟和關閉記憶
為了證實 CREB 在記憶分配中的作用,我們轉向了近年來改變了記憶研究的新開發的方法。這些實驗室技術使得啟用或關閉神經元成為可能——實際上,可以引發或沉默記憶。
舉一個例子,當時在我的實驗室工作的周宇對一小部分小鼠杏仁核神經元進行了基因改造,使它們具有更高的 CREB 水平,並表達了由加利福尼亞州拉霍亞市索爾克生物研究所愛德華·卡拉威實驗室設計的另一種蛋白質。卡拉威的精巧蛋白質使我們能夠在我們選擇的時間沉默 CREB 神經元。當我們關閉具有高 CREB 的神經元時,留下蛋白質水平較低的對應神經元仍然活躍,情緒記憶被抑制,這一結果提供了證據,表明具有較高 CREB 水平的神經元更有可能參與記憶儲存。
我們知道較高水平的 CREB 可以決定哪些細胞儲存給定的記憶,但我們不知道這是如何發生的。斯坦福大學的羅伯特·馬倫卡和他的同事發現,增加某些神經元中的 CREB 意味著它們更容易被啟用。這種興奮性的增加是否是具有較高 CREB 水平的神經元被選用於記憶儲存的原因?
為了解決這個問題,周修改了杏仁核神經元以產生更多 CREB。使用微小的微電極,她確定了這些神經元有多容易被啟用,這是興奮性的一種度量。結果證實,與未改變的對應神經元相比,修改後的神經元更容易被開啟。興奮性的升高(增強了接收和傳遞在神經元之間傳遞資訊的電脈衝的準備狀態)表明,細胞可能更好地準備好啟動記憶形成所需的一系列過程。
為了檢驗這個想法,周還研究了涉及具有更多 CREB 的神經元的突觸連線。大量證據表明,突觸連線強度的增加對於記憶形成至關重要。在對小鼠進行一項隨後引發情緒記憶的任務訓練後,她測試了杏仁核神經元的突觸連線強度,這些神經元具有更高的 CREB 水平,看看它們是否比未被改變以產生更多 CREB 的細胞具有更強的連線。
為了做到這一點,她用微小的電流刺激這些細胞的突觸,並用嵌入細胞內的微小電極記錄它們的反應。正如預期的那樣,具有較高 CREB 的杏仁核神經元比其他細胞具有更強的突觸,這一結果與它們更有可能儲存情緒記憶的想法一致。
在更新近的工作中,喬斯林的實驗室證明,透過對一組預先確定的杏仁核神經元進行基因工程改造,使其具有一種特定型別的離子通道,可以增加這些神經元的興奮性,從而將恐懼體驗的記憶儲存在其中。離子通道在細胞表面形成孔隙,喬斯林選擇的特定離子通道允許這些細胞更容易被啟用。同樣,霍華德·休斯醫學研究所珍妮莉亞研究園區(位於弗吉尼亞州阿什本)的神經科學家阿爾伯特·李的實驗室報告說,當動物在跑道上跑動時,人為地增加特定位置的海馬神經元的興奮性,使這些神經元更有可能對跑道上的那個位置做出反應,這一結果與我們的發現一致,即興奮性在決定哪些細胞參與儲存給定記憶中起著至關重要的作用。
最後,我們的小組以及喬斯林的小組都利用了一項突破性技術,稱為光遺傳學,該技術使用光來啟用或抑制神經元。我們使用該技術來開啟具有較高 CREB 水平的特定神經元。托馬斯·羅傑森和巴拉吉·賈亞普拉卡什,當時都在我的實驗室工作,他們首先對杏仁核神經元進行基因工程改造,以產生更多 CREB 和視紫紅質通道蛋白 2 (ChR2),這是一種由藍光啟用的離子通道。然後我們表明,當我們使用光開啟具有較高 CREB 的杏仁核神經元時,我們可以人為地觸發小鼠對恐懼記憶的回憶,但當開啟蛋白質水平較低的神經元時則不行,這證實了記憶儲存在這些神經元中。
連線起來
2009 年,我被要求寫一篇關於我們記憶研究的文章,我藉此機會介紹了我們關於記憶如何隨時間推移而聯絡起來的想法。CREB 調節哪些細胞形成給定記憶的能力——換句話說,記憶分配——使我提出了一個假設,即這個過程可能是連線不同記憶的關鍵,我的實驗室現在稱之為“分配以連線”假說。由於記憶分配發生在具有更高 CREB 且更容易啟用的神經元子集中,因此這個過程為這些神經元準備好儲存另一個記憶。當兩個記憶共享許多相同的神經元時,它們就會正式連線起來。
因此,在回憶兩個記憶中的一個時啟用這些神經元會觸發對另一個記憶的回憶。這個想法的關鍵預測是,在一天之內形成的、時間上更接近的兩個記憶比間隔較長時間的記憶更有可能被連線起來。如果間隔遠大於一天,第二個記憶不再受益於第一個記憶觸發的興奮性,因此儲存在不同的神經元群體中。記憶連線的時間限制性質是有道理的,因為一天之內發生的事件比相隔一週的事件更有可能彼此相關。
撰寫文章和概述這些想法讓我更加投入到我們如何檢驗它們的挑戰中。“分配以連線”假說很簡單,但我們如何證實其合法性根本不清楚。測試不得不等待合適的時機。
當丹尼斯·J·蔡和賈斯汀·肖伯(當時都在我的實驗室工作)加入該專案時,情況發生了變化。蔡提出了一個聰明的想法。她與肖伯一起,在同一天內、在五個小時的間隔內,將小鼠暴露在兩個房間中,希望這兩個房間的記憶能夠聯絡起來。後來,她在第二個房間裡給了它們輕微的爪子電擊。正如預期的那樣,當她隨後將小鼠放入它們受到電擊的房間時,它們僵住了,大概是因為它們記得在那裡受到了電擊。小鼠會僵住作為對恐懼的自然反應,因為大多數捕食者在獵物移動時更容易注意到它們。
當蔡和肖伯將小鼠放入中性房間時,關鍵結果出現了。我們推斷,如果兩個房間的記憶聯絡在一起,中性空間中的小鼠會想起在另一個房間裡受到電擊,因此會預感到恐懼而僵住——這正是我們發現的。
我們還猜測,如果兩個記憶間隔七天,它們就不太可能被連線起來。事實上,在較長時間間隔後,再次將動物暴露在中性房間中並沒有讓它們想起電擊室,它們也沒有僵住。一般來說,對於遠大於一天的時間間隔,記憶仍然是未連線的。值得注意的是,加州大學聖地亞哥分校吉爾·魯特格布實驗室的研究表明,我們實驗中使用的腦區非常能夠區分我們在測試的時間框架內(數小時)的時間間隔,這一結果與我們的發現相符。
這些行為發現令人興奮,但它們並沒有檢驗該假說的關鍵預測——在緊密間隔形成的不同記憶儲存在同一大腦區域的重疊神經元群體中。這種物理重疊將兩個記憶聯絡起來,因此對一個記憶的回憶會讓人想起另一個記憶。
視覺化記憶
要真正檢驗“分配以連線”假說,就需要在大腦中看到記憶被創造出來的過程。用於對活體小鼠的神經元進行成像的技術已經在使用中,但它們都需要將小鼠的頭部固定在大型顯微鏡上,這種設定不利於檢驗該假說所需的行為實驗。
不過,我發現令人驚奇的是,在我的職業生涯中,正確的技術經常在我們最需要它的時候出現。我碰巧參加了加州大學洛杉磯分校舉辦的研討會,斯坦福大學的馬克·施尼策在研討會上介紹了他實驗室剛剛發明的一種微型顯微鏡,它可以視覺化自由活動小鼠的神經元活動。這種兩到三克重的顯微鏡可以像帽子一樣安裝在動物的頭上。該儀器正是我們小組追蹤給定記憶啟用的神經元所需的。施尼策和他的同事已經表明,它可以用來跨時間追蹤我們正在研究的神經元。因此,我們可以使用他們革命性的微型顯微鏡來檢驗我們的“分配以連線”假說的關鍵預測:在一個記憶期間活躍的許多神經元在幾個小時後建立另一個連線記憶期間再次活躍起來。
我們對這項奇妙發明的承諾感到非常興奮,因此我們決定設計我們自己的顯微鏡版本。我們與加州大學洛杉磯分校的佩曼·戈爾沙尼和巴爾吉特·卡克的實驗室合作,共同聘請了一位才華橫溢的博士後研究員丹尼爾·阿哈羅尼,他繼續設計了我們後來稱之為加州大學洛杉磯分校迷你顯微鏡的東西。與施尼策顯微鏡類似,我們的迷你顯微鏡配備了一個鏡頭,可以嵌入在我們想要記錄的腦細胞附近。該裝置卡在固定在動物頭骨上的底板上,在訓練任務和記憶測試期間保持穩定。正如我們從其他研究人員那裡借用技術一樣,我們也樂於分享。我們是科學界開源運動的積極支持者,並將我們用於加州大學洛杉磯分校迷你顯微鏡的設計和軟體提供給了全球數百個其他研究小組。
為了視覺化迷你顯微鏡的神經元活動,蔡和她的同事特里斯坦·舒曼利用了一種成像技術,該技術對動物的神經元進行基因工程改造,使其在細胞中的鈣水平升高時發出熒光——這被稱為基因編碼鈣指示劑。
我們決定專注於海馬體的 CA1 區域,因為它在學習和記憶地點(例如我們在行為實驗中使用的房間)方面發揮著作用。戴著迷你顯微鏡帽子的小鼠被放置在兩個房間中。我們想知道暴露於不同房間之間的時間間隔是否會影響哪些神經元被啟用。
圖片來源:塔米·托爾帕
結果超出了我們的預期!基本上,我們的迷你顯微鏡和行為實驗表明,當小鼠連線兩個房間的記憶時,當動物訪問第一個房間時變得活躍的許多 CA1 神經元在它們在第二個房間探索時也被開啟。如果訪問之間的間隔約為五個小時,小鼠會在相似的神經元簇中形成兩個記憶。當時間間隔增加到七天時,這種重疊的啟用模式沒有出現。
我們對這個發現感到高興,因為它證實了“分配以連線”假說的基本前提:當記憶儲存在重疊的神經元群體中時,它們會耦合。如果您稍後重新啟用為兩個記憶中的任何一個形成的神經元集合,它會刺激另一個記憶並促進其回憶。
標記記憶
為了進一步驗證迷你顯微鏡的結果,蔡轉向了神經科學家馬克·梅福德(現任斯克裡普斯研究所)開發的另一種方法。這項實驗涉及梅福德的技術,稱為 TetTag 系統(代表四環素標記)。當轉基因小鼠訪問房間時形成記憶時,TetTag 會用熒游標記標記啟用的神經元,該標記可保持完整數週。
然後,對動物的屍檢研究可以比較最近啟用的神經元——用記憶形成後立即表達的基因標記——與用長期標記標記的神經元。此步驟不僅識別出由一個事件開啟的神經元——在這種情況下,一個神經元具有單個熒游標記——還識別出由兩個事件啟用的神經元:兩個標記都發出熒光。
蔡和她的團隊使用與以前相同的實驗設定表明,在短短的五個小時間隔內,每個具有雙重標記的記憶的編碼神經元之間的重疊明顯大於偶然預期的重疊。對於七天的時間間隔,兩次體驗之間的重疊沒有明顯高於偶然水平。
喬斯林的加拿大多倫多研究小組進行的其他實驗提供了更多證據來證明我們的記憶連線假說的有效性。她的研究小組不僅進行了不同版本的神經元標記實驗,科學家們還發現了記憶連線的獨立行為證據。多倫多研究人員推斷,如果編碼兩個記憶的神經元群體重疊,第一個記憶觸發的 CREB 水平的增加也會加強第二個記憶。但是,喬斯林的團隊沒有像我們的工作那樣將小鼠暴露於不同的地點,而是訓練動物學習識別兩種不同的音調。如果兩個訓練課程發生在六小時內,而不是六到二十四小時內,則對第一個音調的訓練會加強對第二個音調的記憶。
最近,日本富山大學的豬口薰和他的同事們將這項分析更進一步。他們使用光遺傳學來滅活兩個不同情緒記憶共享的細胞群,同時保持其他細胞不受干擾,包括那些對每個記憶都是唯一的細胞。研究人員表明,透過滅活共享細胞,他們能夠破壞兩個記憶之間的連線,而不會影響對每個個體記憶的回憶。這項優雅的實驗直接證明了兩個記憶共享的神經元是記憶連線的關鍵。它也增加了為我們初步的“分配以連線”假說提供獨立證據的實驗室數量。
改善衰老過程中的記憶
接下來,我們決定研究老年小鼠的記憶連線。與年輕小鼠相比,老年小鼠大腦(包括海馬體 CA1 區域的神經元)中的 CREB 水平較低,因此興奮性也較低。瞭解了這一點,我們預測老年小鼠在連線記憶方面應該會遇到困難。因此,蔡和她的同事著手重複我們在老年動物身上已經完成的許多相同實驗。結果讓我們感到驚訝。經驗豐富的科學家知道,假說只是工具。我們不期望它們一定是正確的。不可避免的失敗幫助我們沿途重塑我們的想法。但這一次,我們的預感被證明是正確的。
.png?w=2000)
圖片來源:塔米·托爾帕(插圖);來源:“共享神經元集合連線時間上接近的不同情境記憶”,丹尼斯·J·蔡等,《自然》,第 534 卷;2016 年 6 月 2 日(神經元影像)
我仍然記得蔡氣喘吁吁地衝進我的辦公室。她告訴我,中年小鼠儘管記得每個房間,但確實在連線記憶方面存在問題,即使它們在相隔五小時的時間內暴露於房間中,這個間隔對於年輕小鼠來說沒有困難。與年輕成年小鼠相比,老年動物的迷你顯微鏡成像顯示儲存記憶之間缺乏重疊。
我們既興奮又持懷疑態度,因此我們立即回去重複了實驗。第二次,結果變得更加令人信服。CREB 水平較低的中年小鼠的神經元不如年輕小鼠的神經元容易連線記憶。
這些結果鼓舞我們擴大研究範圍。我們能否在老年小鼠探索兩個房間時,人為地增加 CA1 神經元子集的興奮性,確保在一個房間中啟用的一些 CA1 神經元在動物移動到第二個房間時也被開啟?
為了實現這一點,我們利用了一種突破性的方法,該方法在細胞表面對受體進行基因工程改造,從而可以控制細胞的功能。該技術有一個令人難忘的技術縮寫詞 DREADD(代表設計師受體專門被設計師藥物啟用)。啟用 DREADD 受體使我們能夠在動物探索兩個房間時開啟同一組 CA1 神經元,從而在它們對兩個圍欄的記憶之間建立聯絡。
我必須承認,起初這個實驗的想法聽起來很荒謬。有很多原因可能導致它失敗。首先,對地點的記憶涉及數百萬個神經元,這些神經元分佈在多個相互連線的大腦區域,而不僅僅是 CA1 區域。衰老可能會影響許多(如果不是全部)這些區域的記憶連線過程。因此,即使我們成功地增加了 CA1 神經元子集的興奮性,這些細胞也可能不是正確的細胞。更重要的是,我們可能沒有觸發正確水平的興奮性。
但實驗成功了。這種“孤注一擲”試驗的關鍵在於平衡時間和金錢的投入與可能帶來的潛在回報。然而,在這種情況下,我可以肯定地說,幸運之神站在我們這邊。透過恢復中年小鼠特定 CA1 神經元子集中興奮性的增加,我們能夠將兩個記憶分配給許多相同的 CA1 神經元,從而恢復這些中年小鼠的記憶連線。
來自齧齒動物和人類的其他實驗室的研究也闡明瞭一個記憶如何與另一個記憶交織在一起。波士頓大學已故神經科學家霍華德·艾興鮑姆證明,大鼠能夠找到共享內容的記憶之間的聯絡。德克薩斯大學奧斯汀分校的神經科學家艾莉森·普雷斯頓和她的同事表明,當記憶共享內容時,人類可以更容易地將它們聯絡起來。回憶起一個可能會帶回另一個。2017 年,同一研究小組還表明,時間上的接近性促進了跨時間的記憶整合,這一結果在人類身上與我們在小鼠身上的結果驚人地相似。
我們可支配的用於測量和控制神經活動的工具庫不斷增長,這開始揭示我們大腦用來組織資訊的機制。我們的團隊現在正試圖以新的方式擴充套件這項工作。我們正在與希臘研究與技術基金會分子生物學與生物技術研究所的計算神經科學家帕納約塔·波拉齊合作,構建計算機模型來模擬記憶如何以及何時連線起來。我們也在試圖弄清楚控制不同大腦結構中記憶連線所需時間間隔的機制。
到目前為止,多個實驗室進行的大量廣泛實驗都強烈支援“分配以連線”假說。我們希望對記憶如何糾纏在一起的理解可能有助於我們開發治療記憶問題的療法,這些記憶問題在廣泛的精神疾病中都很常見,從與年齡相關的認知衰退到精神分裂症、抑鬱症和雙相情感障礙。除了臨床意義之外,我們描述的研究還反映了記憶研究中一個激動人心的新時代,在這個時代,我們所做的實驗不再受我們擁有的技術的限制,而是受我們想象力的範圍的限制。