
一只孤獨的霸王龍 嗅著白堊紀潮溼的空氣,嗅到了樹林那邊一群三角龍正在吃草的氣味。當這隻掠食者掃描洪泛區時,它的視力突然聚焦。一隻三角龍脫離了獸群,遊蕩到了襲擊範圍之內。霸王龍靜靜地站著,制定了一個攻擊計劃,預測它必須以精確的角度與目標相交,才能在三角龍重新回到獸群的安全地帶之前發起攻擊。下午的寂靜被打破了,掠食者衝破森林邊緣的低矮樹枝,開始了猛烈的追擊。
霸王龍在如此多的書籍、遊戲和電影中獵殺三角龍,以至於這種遭遇已經變得老生常談。但是,這樣的場景真的在現實生活中發生過嗎?霸王龍會透過視覺還是嗅覺來識別獵物?三角龍會被響亮的樹枝斷裂聲警告,還是因為無法定位聲音來源而依然渾然不覺?霸王龍會像貓一樣計劃攻擊,還是會像鯊魚一樣不分青紅皂白地猛撲過去?
自從恐龍在19世紀早期首次被描述以來,古生物學家一直在爭論它們的智力、感官能力和行為複雜性。早期的研究依賴於天然顱腔內模,這是當沉積物填充頭骨中的空隙時形成的鑄型。這些鑄型複製了生物體內的顱腔內容物的形狀。長期以來的傳統觀點認為,所有恐龍的大腦都很小,因此行為也很簡單。關於恐龍智力最有趣的例子可能來自19世紀的古生物學家奧瑟內爾·查爾斯·馬什,他假設裝甲恐龍劍龍在臀部附近有一個第二個大腦,以補充其頭骨中核桃大小的大腦。這個想法是基於恐龍骨盆附近脊髓管的模糊的顱腔形狀的擴張。這種神秘的擴張現在被認為是糖原體——一種儲存富含能量的葡萄糖的結構,在一些現代鳥類中也存在於相似的位置。
支援科學新聞報道
如果您喜歡這篇文章,請考慮支援我們屢獲殊榮的新聞報道,方式是 訂閱。透過購買訂閱,您正在幫助確保未來能夠繼續報道關於塑造我們今天世界的發現和想法的具有影響力的故事。
當今的古生物學家仍然不相信劍龍有能力進行更高層次的推理。但在最近幾年,科學家們對其他一些恐龍,特別是那些產生鳥類的獸腳亞目譜系的成員的認知能力的評價有所提高。隨著微型計算機斷層掃描 (CT) 掃描等新技術的出現,我們現在可以重建大腦的體積和表面地形,而不必完全依賴於稀有的天然顱腔內模,這大大擴充套件了可供研究的物種數量。先進的成像技術還在教我們恐龍可能如何使用它們的大腦。我們現在擁有了回答問題的工具,即早已消失的動物如何感知周圍的世界,以及在恐龍時代掠食者與獵物相遇時到底發生了什麼。
霸王龍在遲鈍的劍龍和會使用工具的烏鴉之間的智力譜系中處於什麼位置?在去年秋天發表的一篇備受矚目的論文中,範德比爾特大學的神經科學家蘇珊娜·赫庫拉諾-霍澤爾認為,霸王龍的智力大約相當於狒狒——這是一個令人震驚的結論,因為靈長類動物,以其龐大的大腦,是一些最聰明的動物。我們花了很長時間思考大腦體積如何隨身體大小縮放,以及這種關係對於已滅絕的恐龍和鳥類的大腦功能意味著什麼,我們很感興趣地看到了關於這項研究的頭條新聞。從表面上看,暴君蜥蜴王的大腦與它的身體大小相比顯得相當渺小。這種六噸重恐龍的大腦重量不到一磅,與非洲象的11磅大腦相比顯得微不足道,儘管非洲象是現存最大的陸地哺乳動物,但它的身體比霸王龍小。
赫庫拉諾-霍澤爾認為,當涉及到智力時,大腦大小和身體大小之間的關係並不重要。她說,重要的是端腦中神經元的原始數量,端腦是大腦前部的一個區域,不僅包括處理氣味的嗅球,還包括大腦皮層,更高層次的認知功能(如決策)發生在這裡。科學家們之前對脊椎動物大腦中存在多少神經元只有粗略的瞭解,因為在不同的物種中,它們在大腦的不同部位的密度可能更高或更低。
擁有靈長類動物智力的霸王龍將是可怕的。然而,我們認為應該有一些注意事項。
赫庫拉諾-霍澤爾和里約熱內盧聯邦大學的羅伯託·倫特發明了一種計算神經元的技術,稱為各向同性分餾器法。它使用特殊的化學物質來溶解大腦,基本上是製作腦湯。一種熒光染料會給神經元的細胞核染色,使其發光並易於觀察。研究人員可以精確地計算出湯中少量均勻樣本中發光的細胞核,然後推斷出活體大腦中神經元的總數。使用這種方法,赫庫拉諾-霍澤爾和她的同事計算出人腦大約有1000億個神經元,證實了早期的估計。
各向同性分餾器法很巧妙,但科學家們永遠不會有實際的霸王龍大腦來溶解。相反,赫庫拉諾-霍澤爾依賴於現存溫血和冷血物種的端腦大小和神經元數量之間的比例關係,根據已滅絕恐龍的推斷代謝率,將它們代入兩個方程中的一個。這種比例關係在脊椎動物中差異很大。冷血或外溫物種的神經元密度往往低於溫血或內溫物種。
例如,各向同性分餾器資料表明,一隻159磅重的尼羅鱷大約有8150萬個神經元,而一隻73磅重的鴯鶓大約有13億個神經元——幾乎是鱷魚的16倍,儘管只有它一半的大小。赫庫拉諾-霍澤爾認為,大多數獸腳亞目恐龍可能是內溫動物,因此接近神經元密度連續體的鳥類端。在這種假設下,一個重約12盎司的霸王龍端腦將包含大約30億個神經元,與許多靈長類動物相當。

梅薩·舒馬赫
擁有靈長類動物智力的霸王龍將是可怕的。然而,我們認為應該有一些注意事項。除了獸腳亞目恐龍是內溫動物的合理推測之外,30億個神經元的估計還依賴於其他一些假設。其中之一是整個顱腔都被大腦佔據,這顯然與霸王龍顱腔內模的形態不符。化石證據表明,在許多恐龍中,鼻竇和血管等結構佔據了顱腔的很大一部分。實際的大腦體積將小於原始的顱腔體積。事實上,基於現代鱷魚的研究表明,霸王龍的大腦可能只佔整個顱腔體積的30%。
同樣重要的是要注意,不同的腦區具有不同的功能。有些腦區致力於呼吸調節等基本任務,而另一些腦區則支援語言等複雜功能。如果大腦皮層在一個物種的大腦大小中占主導地位,而擴大的嗅球在另一個物種的大腦大小中占主導地位,那麼具有相同端腦大小的兩個物種可能具有截然不同的認知能力。對於現存物種,可以使用細胞染色或磁共振成像等技術來確定不同腦區之間的邊界。
霸王龍真的能夠嗅到風,並在看到獵物或腐肉之前很久就識別出它們,以便進行清掃。
對於恐龍來說,定義這些邊界要困難得多,因為我們所能利用的只是表面拓撲結構。我們中的一位(巴拉諾夫)花費了她職業生涯的很大一部分時間來繪製骨性地標,這些地標可以更好地估計顱腔內模主要腦區的體積。這項工作表明,大腦皮層的擴張出現在更特化的獸腳亞目恐龍中,如竊蛋龍和馳龍——這些譜系的分支時間比暴龍科晚得多。相比之下,較早分化的獸腳亞目恐龍,如霸王龍,其大腦皮層相對較小,整個端腦的大部分都讓給了嗅球。
一旦我們考慮到顱腔內模中容納的非神經組織體積,霸王龍的端腦中不太可能有任何型別的30億個神經元。我們同意霸王龍 是一個熟練的掠食者,但我們認為它可能不具備靈長類動物身上看到的先進的計劃能力或協調的社會狩獵能力。
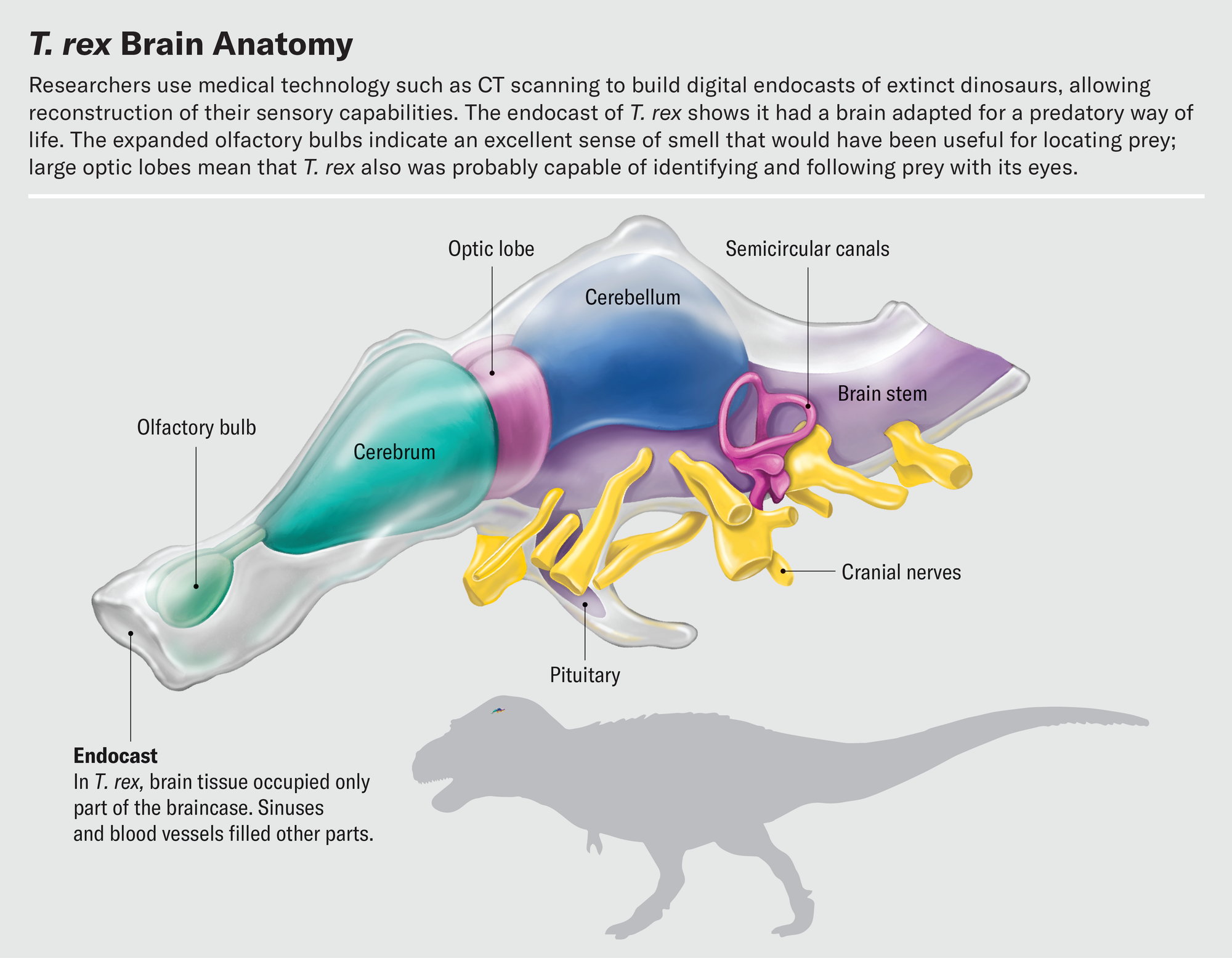
使用CT掃描構建的化石顱腔內模進行研究的最好之處之一是,我們可以研究內部特徵,而不會損壞化石本身。虛擬地切割化石顱腔內模是一種度過下午的愉快方式。訓練有素的古生物學家透過逐片探索大腦切片,可以使用骨性地標來解讀關鍵腦區的邊界,並以數字方式隔離這些區域。當我們從前到後分割大腦時,我們遇到的第一個結構是嗅球。嗅球的形狀在恐龍及其親屬中差異很大。鱷魚的嗅球大約有小葡萄大小,位於通往大腦其餘部分的長柄末端。大多數鳥類的嗅球要小得多;事實上,在許多物種中,它們幾乎無法與大腦皮層的其餘部分割槽分開來。
顧名思義,嗅球促進嗅覺——一種依賴於稱為氣味分子的微小分子的感覺。吸入的氣味分子與鼻組織中的受體結合,鼻組織透過神經元與嗅球進行通訊。令人驚訝的是,每個受體都會產生一種氣味分子-受體蛋白,這種蛋白會根據特定型別的氣味分子進行調整。這些蛋白質中的每一種都由不同的嗅覺受體基因編碼。基因組測序顯示,鳥類擁有182到688個功能性嗅覺受體基因。
在最近的一項研究中,都柏林大學學院的格雷厄姆·休斯和約翰·菲納雷利調查了恐龍的嗅覺。恐龍的嗅覺受體基因數量無法直接測量,但由於嗅球大小與受體數量相關,因此嗅球尺寸可以作為衡量動物探測氣味能力的指標。休斯和菲納雷利發現,總的來說,恐龍的嗅球比鳥類大得多。在獸腳亞目恐龍中,雜食性似鳥龍科動物的嗅球最小,而肉食性物種的嗅球最大。霸王龍名列前茅,其嗅球尺寸與存在600多個嗅覺受體基因的情況相符。這個數字與家貓相當,並且高於幾乎所有現代鳥類。我們假設的跟蹤三角龍的霸王龍真的能夠嗅到風,並在看到獵物或腐肉之前很久就識別出它們,以便進行清掃。
梅薩·舒馬赫;資料來源:勞倫斯·威特默和瑞安·裡奇利在《解剖學記錄》第292卷中發表的“關於暴龍(恐龍,獸腳亞目)大腦、顱腔和耳朵區域的新見解,及其對感官組織和行為的影響”,2009年;以及阿什利·莫哈特在俄亥俄大學2016年博士論文中發表的“粗略解剖腦區近似 (GABRA):評估已滅絕的初龍的大腦大小、結構和進化”(顱腔內模參考文獻)
其他基於CT的顱腔內模研究暗示了霸王龍的眼睛有多麼敏銳。透過虛擬地切割顱腔內模並隔離視葉,我們發現這些結構在已滅絕的獸腳亞目恐龍和現存鳥類中的相對大小相似。因此,鳥類一定是從它們的非鳥類祖先那裡繼承了它們的視覺敏銳度。眾所周知,鳥類是高度視覺化的動物——一隻鷹可以從半英里外發現一隻兔子,一隻飛行中的燕鷗可以追蹤到水面下半英寸長的魚,並從上方將其捕獲。這種對視覺的依賴反映在鳥類大腦的結構中。視葉處理視覺資訊並位於大腦皮層之後,是大腦最突出的特徵之一。通常情況下,一個區域相對於大腦其餘部分越大,該區域對動物就越重要。這對於視葉來說當然是成立的。
我們可以從已滅絕恐龍的進化關係中推斷出它們的一些視覺能力。例如,鳥類和鱷魚(已滅絕恐龍最近的現存親戚)都具有看到顏色的視網膜受體型別。因此,恐龍很可能也具有色覺。然而,視覺是一種複雜的感覺。準確地重建已滅絕恐龍的視覺能力需要我們超越這些型別的推斷。
使猛禽如此擅長捕捉獵物的一個因素是立體視覺——一種增強的感知深度的能力。這種複雜能力背後的感官適應令人驚訝地簡單。它僅僅與眼球的位置有關。眼睛位於頭部側面的動物,如壁虎,缺乏重疊的視野,因此它們的三維視覺不好。眼睛位於頭部前方的動物具有在鼻子前方重疊的視野。在這個重疊的空間內,每隻眼睛從略微不同的角度感知相同的資訊,產生所謂的雙眼視覺。
嘗試將目光固定在離您大約一英尺左右的物體上,然後閉上一隻眼睛,再閉上另一隻眼睛。物體似乎會移動,因為您在鼻子前面看到了相同的影像,但角度不同。大腦整合這些略有不同的影像以產生視覺深度。眼睛橫向定位的動物透過用一隻眼睛看某物,移動頭部,然後用另一隻眼睛看它來判斷深度——這不是一種特別隱秘的技術。對於掠食者來說,雙眼視覺特別有用,因為它允許動物識別並鎖定獵物,而無需透過移動頭部來暴露其位置。
為了確定已滅絕的動物是否具有立體視覺,我們必須考慮它們的眼眶位置,眼眶是頭骨中容納眼球的空間。俄勒岡大學的肯特·史蒂文斯對這個問題採取了一種創造性的方法,他根據幾種獸腳亞目恐龍的骨骼結構雕刻了它們的頭部。從那裡,他能夠繪製出它們的視野範圍,包括任何障礙物,如角或特別大的鼻子。他發現霸王龍的眼睛朝前,鼻子狹窄,不會阻礙其視線,使其視覺敏銳度與鷹相似。與電影《侏羅紀公園》中虛構的霸王龍不同,真正的霸王龍不需要獵物移動才能將其從背景中挑出來。恐爪龍類,如傷齒龍和迅猛龍,可能更擅長識別獵物。由於它們的頭部略微向前傾斜,它們擁有更寬廣的視野和增強的深度感知能力,可能與貓頭鷹媲美。
我們可以從已滅絕動物的眼眶中推斷出比感知深度更多的資訊。這些開口的大小及其相關的骨骼提供了關於它們所容納的眼球大小的線索。較大的眼眶通常表明眼球較大。相對於頭部大小而言,較大的眼球在夜行動物中很常見,因為它們可以容納更多的視網膜感光細胞。此外,包括魚類、一些已滅絕的哺乳動物和爬行動物(包括非鳥類恐龍和鳥類)在內的許多動物的眼球,在其被稱為鞏膜的纖維外層內嵌有骨性或軟骨環。由於骨骼是堅硬的,這些鞏膜環限制了眼球的運動,包括瞳孔可以張開的程度。這會影響到達視網膜感光細胞的光量。較大的鞏膜環允許瞳孔張得更大,從而讓更多的光線進入眼睛。
幾年前,巴拉諾夫曾是南非威特沃特斯蘭德大學的喬納·喬伊尼爾領導的一個團隊的成員,該團隊研究了非鳥類恐龍眼眶中這些環的大小,以確定它們是喜歡在白天還是晚上活動。由於鞏膜環不附著在頭骨的任何部分,它們很容易與骨骼的其餘部分分離,並且很少儲存在化石中。在保留它們的罕見標本中,它們並不總是被發現處於原位。在這項研究中,研究人員使用了具有儲存完好的鞏膜環的已滅絕恐龍的CT影像,以數字方式隔離這些骨骼並在眼眶內重建它們。他們的結論是,它們的眼眶和鞏膜環的比例表明,許多已滅絕的恐龍主要在白天活動。
但是,一種家貓大小的阿瓦雷茲龍科獸腳亞目恐龍,沙漠爪龍,提供了不同的結果。該團隊檢查了在蒙古晚白堊世沙漠沙丘沉積物中發現的一個沙漠爪龍頭骨,並驚訝地發現其適應性與最著名的現存夜行動物之一——穀倉貓頭鷹趨同。穀倉貓頭鷹有大的眼眶和充足的鞏膜環,這使得它們的瞳孔可以張得非常大。這種結構可以讓大量的光線湧入眼睛,在最黑暗的條件下提供影像。在阿瓦雷茲龍科動物中也存在這些相同的特徵表明,它們也是夜間活動的掠食者。

梅薩·舒馬赫
沒有人正式分析過霸王龍的眼眶形態,以深入瞭解這種恐龍最活躍的時間。然而,我們確實知道,霸王龍有一個很大的眼眶,形狀幾乎像一個鑰匙孔。如果眼睛填滿了整個眼眶,那麼我們或許可以推斷出霸王龍是夜間活動的,但來自近緣物種的鞏膜環表明,它的眼球只填滿了眼眶的一小部分——因此可能無法收集足夠的光線在夜間發揮太大作用。事實上,英國伯明翰大學的斯特凡·勞特enschlager領導的頭骨應力數字分析表明,霸王龍的大眼眶有助於分散其強力咬合產生的巨大應力,而不是容納一個大眼睛。
在脊椎動物頭骨的後部附近是一個有趣而複雜的結構:內耳。雖然從技術上講它不是大腦的一部分,但它是一個重要的感覺器官,向大腦傳送大量資訊。兩種特殊感覺,平衡和聽覺,由內耳所謂迷路的不同部分控制。迷路包括半規管、環狀結構,用於檢測頭部的旋轉運動;前庭,一種球狀結構,用於感知前後和左右運動;以及耳蝸,用於感知聲音振動。內耳充滿了液體,並利用該液體內毛細胞的偏轉來檢測這些不同型別的資訊。
2021年,現任職於史密森尼學會的邁克爾·漢森和他的同事對半規管和前庭的形狀進行了複雜的分析,以推斷非鳥類恐龍的主要運動方式。他們製作了內耳空間的虛擬顱腔內模,以估計迷路在生命期間的形態。他們的資料表明,大多數恐龍的活動僅限於在地面上行走和奔跑。但是在導致鳥類的譜系中,耳朵的結構發生了變化。在其他變化中,半規管變長了。這種伸長允許恐龍做出和解讀更復雜的頭部運動。霸王龍沒有長的半規管,這表明它只能行走或奔跑。但是,一些更接近迅猛龍的傷齒龍科恐龍的耳朵可以感知與飛行相關的複雜運動。這些恐龍可能能夠滑翔或進行雛形的飛行,在現代鳥類起飛之前在空中移動。
我們可以從耳蝸中尋找恐龍聽力的線索。耳蝸的長度與聽覺靈敏度相關。較長的耳蝸管允許基底乳突的伸長,基底乳突是容納拾取聲音振動的毛細胞的結構。蜥蜴和烏龜往往具有短而粗壯的耳蝸,最適合檢測低音調的聲音。相比之下,鱷魚和鳥類具有長而細長的耳蝸,擅長檢測高音調的聲音。
許多鳥類會唱出悅耳的歌曲來吸引配偶和保衛領地,因此它們對高音調發聲的敏感性提高似乎是理所當然的。然而,耳蝸的伸長並非起源於鳥類,而是起源於鳥類和鱷魚的共同祖先。奇怪的是,鱷魚的發聲僅限於低音調的閉口咕嚕聲——而不是細長的耳蝸擅長檢測的那種聲音。與其他爬行動物相比,鱷魚為何具有如此先進的耳蝸形狀一直是個謎。
漢森和他的合作者提出了一個巧妙的解釋。他們假設,將鱷魚、已滅絕的恐龍和鳥類與早期分化的爬行動物區分開來的耳蝸伸長與親代撫育的進化有關。與大多數爬行動物不同,鱷魚會照顧它們的幼崽。與大多數幼年爬行動物不同,幼年鱷魚會發出唧唧聲以引起父母的注意。也許鳥類和鱷魚的祖先需要能夠探測高音調的聲音,以便聽到它們的幼崽而不是配偶或競爭對手。
與食草的獵物相比,霸王龍的大腦比例較大,這與現代掠食者相似。
這個假設對鳥鳴的進化具有令人興奮的意義。在這種情景中,幼年恐龍可能會發出唧唧聲以引起注意,但成年恐龍的叫聲僅限於簡單的低音調叫聲。在恐龍向鳥類過渡的過程中,一些譜系將高音調發聲的能力保留到了成年期。耳蝸伸長最初因其在增強親代撫育中的作用而受到自然選擇的青睞,然後作為一種預適應,使歌曲能夠在後來的鳥類中出現。
但是進化並非朝著單一方向發展。透過篩選恐龍資料,漢森和他的同事偶然發現了一種不尋常的短耳蝸,它存在於異特龍(霸王龍的近親)中。這一發現表明,這種大型獸腳亞目恐龍失去了對高音調聲音的敏感性。該團隊推測,異特龍可能沒有照顧它們的幼崽,從而消除了聽到它們發聲的選擇壓力。
耳蝸的形狀也幫助我們瞭解了那些古怪的夜行性阿瓦雷茲龍科動物的生態。巴拉諾夫和她的同事發現,沙漠爪龍的耳蝸非常長,以至於它在頭骨底部下方捲曲。已知夜行性鳥類(如貓頭鷹)具有類似的結構。沙漠爪龍的過度伸長的耳蝸管表明,它的聽力適應於非常高頻的聲音,例如昆蟲發出的聲音。該團隊得出結論,這種小型恐龍在亞洲中部沙漠沙丘的黑暗中漫遊,捕獵這些小型獵物。
人們傾向於將古生物學視為一門基於野外研究的學科,專注於在偏遠沙漠地區度過夏天的浪漫魅力,手持鶴嘴鋤,收集對科學界來說是全新的早已滅絕的動物化石。但是,如今,古生物學家很可能在實驗室中使用生物醫學和神經科學的前沿技術來進行他們最重要的發現。正是這些不同方法的結合使我們能夠重建當霸王龍 遇到三角龍時可能真正發生的事情。
我們自己的研究告訴我們,與現代掠食者相似,霸王龍的大腦比例較大,這與食草的獵物相比。它大腦的很大一部分致力於嗅覺,因此霸王龍可能確實會嗅探空氣以尋找它的下一頓飯,無論是沿著樹林線吃草的活三角龍,還是已經死亡並在陽光下腐爛的三角龍。一旦霸王龍識別出氣味,它就可以用它的立體視覺掃描地平線,尋找任何潛在獵物的跡象。它的眼睛將能夠鎖定那隻毫不在意地在遠離獸群安全地帶的植被叢中覓食的三角龍。
當霸王龍衝過樹林時,它可能會驚嚇到一隻在附近樹枝上築巢的小型傷齒龍科恐龍。憑藉其擴大的內耳迷路提供的增強的運動技能,傷齒龍科恐龍可能會從巢穴中滑翔出來,轉移掠食者對其唧唧叫的幼崽的注意力。三角龍可能因這種騷動而感到不安,停止了平靜的吃草,並返回獸群的安全地帶。這仍然是一個老生常談的故事,但這是一個更具科學依據的故事。